https://pubmed.ncbi.nlm.nih.gov/6940487/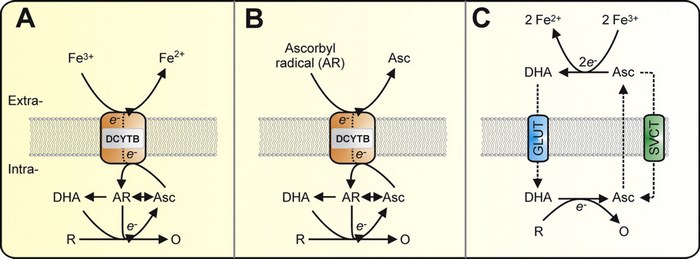 Untitled Document
Untitled Document
Interaction of vitamin C and iron
Food iron is absorbed by the intestinal mucosa from two separate pools of heme and nonheme iron. Heme iron, derived from hemoglobin and myoglobin, is well absorbed and relatively little affected by other foods eaten in the same meal. On the other hand, the absorption of nonheme iron, the major dietary pool, is greatly influenced by meal composition. Ascorbic acid is a powerful enhancer of nonheme iron absorption and can reverse the inhibiting effect of such substances as tea and calcium/phosphate. Its influence may be less pronounced in meals of high iron availability--those containing meat, fish, or poultry. The enhancement of iron absorption from vegetable meals is directly proportional to the quantity of ascorbic acid present. The absorption of soluble inorganic iron added to a meal increases in parallel with the absorption of nonheme iron, but ascorbic acid has a much smaller effect on insoluble iron compounds, such as ferric oxide or ferric hydroxide, which are common food contaminants. Ascorbic acid facilitates iron absorption by forming a chelate with ferric iron at acid pH that remains soluble at the alkaline pH of the duodenum. High cost and instability during food storage are the major obstacles to using ascorbic acid in programs designed to combat nutritional iron deficiency anemia.
Full text
Link:https://pubmed.ncbi.nlm.nih.gov/25048971/
The active role of vitamin C in mammalian iron metabolism: much more than just enhanced iron absorption!
Ascorbate is a cofactor in numerous metabolic reactions. Humans cannot synthesize ascorbate owing to inactivation of the gene encoding the enzyme l-gulono-γ-lactone oxidase, which is essential for ascorbate synthesis. Accumulating evidence strongly suggests that in addition to the known ability of dietary ascorbate to enhance nonheme iron absorption in the gut, ascorbate within mammalian systems can regulate cellular iron uptake and metabolism. Ascorbate modulates iron metabolism by stimulating ferritin synthesis, inhibiting lysosomal ferritin degradation, and decreasing cellular iron efflux. Furthermore, ascorbate cycling across the plasma membrane is responsible for ascorbate-stimulated iron uptake from low-molecular-weight iron-citrate complexes, which are prominent in the plasma of individuals with iron-overload disorders. Importantly, this iron-uptake pathway is of particular relevance to astrocyte brain iron metabolism and tissue iron loading in disorders such as hereditary hemochromatosis and β-thalassemia. Recent evidence also indicates that ascorbate is a novel modulator of the classical transferrin-iron uptake pathway, which provides almost all iron for cellular demands and erythropoiesis under physiological conditions. Ascorbate acts to stimulate transferrin-dependent iron uptake by an intracellular reductive mechanism, strongly suggesting that it may act to stimulate iron mobilization from the endosome. The ability of ascorbate to regulate transferrin iron uptake could help explain the metabolic defect that contributes to ascorbate-deficiency-induced anemia.
Full text
Link:https://pubmed.ncbi.nlm.nih.gov/15743017/
Enhancers of iron absorption: ascorbic acid and other organic acids
Ascorbic acid (AA), with its reducing and chelating properties, is the most efficient enhancer of non-heme iron absorption when its stability in the food vehicle is ensured. The number of studies investigating the effect of AA on ferrous sulfate absorption far outweighs that of other iron fortificants. The promotion of iron absorption in the presence of AA is more pronounced in meals containing inhibitors of iron absorption. Meals containing low to medium levels of inhibitors require the addition of AA at a molar ratio of 2:1 (e.g., 20 mg AA: 3 mg iron). To promote absorption in the presence of high levels of inhibitors, AA needs to be added at a molar ratio in excess of 4:1, which may be impractical. The effectiveness of AA in promoting absorption from less soluble compounds, such as ferrous fumarate and elemental iron, requires further investigation. The instability of AA during food processing, storage, and cooking, and the possibility of unwanted sensory changes limits the number of suitable food vehicles for AA, whether used as vitamin fortificant or as an iron enhancer. Suitable vehicles include dry-blended foods, such as complementary, precooked cereal-based infant foods, powdered milk, and other dry beverage products made for reconstitution that are packaged, stored, and prepared in a way that maximizes retention of this vitamin. The consumption of natural sources of Vitamin C (fruits and vegetables) with iron-fortified dry blended foods is also recommended. Encapsulation can mitigate some of the AA losses during processing and storage, but these interventions will also add cost. In addition, the bioavailability of encapsulated iron in the presence/absence of AA will need careful assessment in human clinical trials. The long-term effect of high AA intake on iron status may be less than predicted from single meal studies. The hypothesis that an overall increase of dietary AA intake, or fortification of some foods commonly consumed with the main meal with AA alone, may be as effective as the fortification of the same food vehicle with AA and iron, merits further investigation. This must involve the consideration of practicalities of implementation. To date, programs based on iron and AA fortification of infant formulas and cow's milk provide the strongest evidence for the efficacy of AA fortification. Present results suggest that the effect of organic acids, as measured by in vitro and in vivo methods, is dependent on the source of iron, the type and concentration of organic acid, pH, processing methods, and the food matrix. The iron absorption-enhancing effect of AA is more potent than that of other organic acids due to its ability to reduce ferric to ferrous iron. Based on the limited data available, other organic acids may only be effective at ratios of acid to iron in excess of 100 molar. This would translate into the minimum presence/addition of 1 g citric acid to a meal containing 3 mg iron. Further characterization of the effectiveness of various organic acids in promoting iron absorption is required, in particular with respect to the optimal molar ratio of organic acid to iron, and associated feasibility for food application purposes. The suggested amount of any organic acid required to produce a nutritional benefit will result in unwanted organoleptic changes in most foods, thus limiting its application to a small number of food vehicles (e.g., condiments, beverages). However, fermented foods that already contain high levels of organic acid may be suitable iron fortification vehicles.
Full text
Untitled Document
Effectiveness of Dietary Intervention with Iron and Vitamin C Administered Separately in Improving Iron Status in Young Women
In order to maintain an adequate iron status in young women, effective dietary interventions should provide sufficient amounts of iron in everyday meals and improve the bioavailability of non-heme iron by providing vitamin C. While some intervention studies administering products rich in vitamin C in conjunction with iron sources showed improved iron status, it is unknown whether a separate administration of products rich in iron and vitamin C may be a successful strategy as well. The aim of this study was to assess the effectiveness of dietary intervention with iron and vitamin C administered separately in improving iron status in young women to prevent iron deficiency anemia. The study was conducted in a group of 29 women aged 18-30, and an 8-week dietary intervention was performed. Study participants with an adequate iron status received 50 g of iron-fortified oat flakes (as a source of non-heme iron) with breakfast and 200 mL of orange juice (as a source of vitamin C) in the second part of the day. Iron status was analyzed based on red blood cells, hemoglobin, hematocrit, serum ferritin, and serum iron, and it was assessed at baseline, after 4 weeks, and after 8 weeks of the intervention. The intakes of iron, vitamin C, and folate were controlled throughout the study period, and menstrual blood loss was estimated. After 8 weeks of intervention, statistically significant differences compared with baseline were observed only for hematocrit, as its level after 8 weeks of intervention was higher than the baseline (p = 0.0491). Comparing subsamples within the dietary intervention considered effective and ineffective for red blood cell levels, it was indicated that lower baseline vitamin C intake may result in a more effective dietary intervention (p = 0.0231). Comparing subsamples within the dietary intervention considered effective and ineffective for hemoglobin, hematocrit, iron, and serum ferritin levels, it was indicated that higher baseline levels of hemoglobin (p = 0.0143), hematocrit (p = 0.0497), iron (p = 0.0101), and serum ferritin (p = 0.0343) respectively may result in a more effective dietary intervention. It was concluded that dietary intervention with iron and vitamin C administered separately may be effective in improving iron status in young women to prevent iron deficiency anemia. It may be concluded that in the studied group, a better baseline iron status and lower baseline vitamin C intake may result in a more effective dietary intervention with iron and vitamin C administered separately to improve iron status in young women.
Full text

